| Part of the Biology series on |
| Evolution |
 |
| Mechanisms and processes |
|
Adaptation |
| Research and history |
|
Evidence |
| Evolutionary biology fields |
|
Ecological genetics |
| Biology Portal · |

{kind=link}
In 1832, while traveling on the Beagle, naturalist Charles Darwin collected giant fossils in South America. On his return, he was informed in 1837 by Richard Owen that fragments of armor were from the gigantic extinct glyptodon, creatures related to the modern armadillos he had seen living nearby. The similarities between these two unusually scaly mammals and their geographic distribution provided Darwin with a clue that helped him develop his theory of how evolution occurs.[1]
Evolution is the biological process in which inherited traits become more or less common in a population over successive generations. Over time, this process can lead to speciation, the development of new species from existing ones. All extant organisms are related by common descent, having evolved over billions of years of cumulative genetic changes from a single ancestor.[2][3]
The basic mechanisms that produce evolutionary change are natural selection and genetic drift; these two mechanisms act on the genetic variation caused by mutation, genetic recombination and gene flow. Through genetic drift the frequency of heritable traits changes randomly and through natural selection organisms with traits that help them to survive and reproduce will have more offspring passing those traits on to the next generation. Thus, more of the offspring will have advantageous traits and fewer will have disadvantageous traits. [2][4][5] Given enough time, this passive process can result in varied adaptations to changing environmental conditions.[6]
An outline of the theory of natural selection was jointly presented to the Linnean Society of London in 1858 in separate papers by Charles Darwin and Alfred Russel Wallace. Detailed support for the theory was then set out in Darwin’s 1859 book, On the Origin of Species. In the 1930s, Darwinian natural selection was combined with the theory of Mendelian heredity to form the modern evolutionary synthesis, also known as "Neo-Darwinism". The modern synthesis describes evolution as a change in the frequency of different versions of genes, known as alleles, within a population from one generation to the next.[6] With its enormous explanatory and predictive power, this theory has become the central organizing principle of modern biology, relating directly to topics such as the origin of antibiotic resistance in bacteria, eusociality in insects, and the biodiversity of Earth's ecosystem.[7][8][9]
Basic processes
Evolution consists of three basic types of processes: those that introduce new variation into a population, those that affect the frequencies of existing genes, and those that induce cooperation into higher order entities (such as genomes, cells, organisms and more complex cooperative systems). Paleontologist Stephen J. Gould once phrased this succinctly as "variation proposes and selection disposes."[10] In natural populations, there is a certain amount of phenotypic variation (meaning differences in traits such as physical appearance, physiology or behavior). This phenotypic variation is the result of variants in gene sequences among the individuals of a population and the interaction of a genotype with the environment, or phenotypic plasticity. There may be one or more functional variants of a gene or locus, and these variants are called alleles. Most sites in the genome (complete DNA sequence) of a species are identical in all individuals in the population; sites with more than one allele are called polymorphic or segregating sites.
Variation
Genetic variation is often the result of a new mutation in a single individual; in subsequent generations the frequency of that variant may fluctuate in the population, becoming more or less prevalent relative to other alleles at the site. This change in allele frequency is the commonly accepted definition of evolution, and all evolutionary forces act by driving allele frequency in one direction or another. Variation disappears when it reaches the point of fixation — a frequency of one, where it replaces the ancestral allele entirely — or when it reaches a frequency of zero and disappears from the population.
Variation is also produced during the production of gametes and the union of gametes at fertilization to produce a zygote (genetic recombination produces variation). In some organisms (like bacteria and plants) the lateral transfer of genetic material or Horizontal gene transfer plays a significant role, and the mixing of genetic material by hybridization (mixing species) produces significant variation (as occurs in plants and in birds).
Heredity
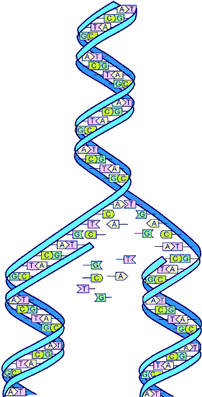
{kind=link}
A section of a model of a DNA molecule.
Gregor Mendel's work provided the first firm basis to the idea that inheritance of traits occurred by means of discrete, particulate units (as opposed to the blending of fluids). He noticed several traits in peas that occur in only one of two clearly discernable forms (e.g., the peas were either "round" or "wrinkled"), and was able to show that the traits were: heritable (passed from parent to offspring); discrete (i.e., if one parent had round peas and the other wrinkled, the progeny were not intermediate, but either round or wrinkled); and were distributed to progeny in a well-defined and predictable manner (Mendelian inheritance). His research laid the foundation for the concept of discrete heritable factors, known today as genes. After Mendel's work was "rediscovered" in 1900, it was discovered that the concepts could have wide applicability, and that most complex traits were polygenetic and not controlled by single unit characters.
Later research gave a physical basis to the notion of genes, and eventually identified DNA as the genetic material, and identified genes as discrete linear elements within DNA. DNA is not perfectly copied, and rare mistakes (mutations) in genes can affect traits that the genes control (e.g., pea shape).
A gene can have modifications such as DNA methylation, which do not change the nucleotide sequence of a gene, but do result in the epigenetic inheritance of a change in the expression of that gene in a trait. Another epigenetic mechanism is via micro RNA's, and RNA interference which serve regulatory roles in gene transcription and translation.
Non-DNA based forms of heritable variation exist, such as transmission of the secondary structures of prions or structural inheritance of patterns in the rows of cilia in protozoans such as Paramecium[11] and Tetrahymena. Investigations continue into whether these mechanisms allow for the production of specific beneficial heritable variation in response to environmental signals. If this were shown to be the case, then some instances of evolution would lie outside of the typical Darwinian framework, which avoids any connection between environmental signals and the production of heritable variation. However, the processes that produce these variations are rather rare, often reversible, and leave the genetic information intact.
Mutation
{kind=link}
Mutation can occur because of "copy errors" during DNA replication.
Genetic variation arises due to mutations that occur in the genomes of all organisms. These mutations are generally considered to be random (with respect to fitness) and to occur at a fairly constant rate in the absence of additional factors. Mutations are permanent, transmissible changes to the genetic material (usually DNA or RNA) of a cell, and can be caused by: "copying errors" in the genetic material during cell division; by exposure to radiation, chemicals, or viruses. In multicellular organisms, mutations can be subdivided into germline mutations that occur in the gametes and thus can be passed on to progeny, and somatic mutations that can lead to the malfunction (or death) of one or more somatic cells (it is possible for multiple somatic mutations to produce cancer).
Mutations that are not affected by natural selection under the prevailing environment are called neutral mutations. The frequency of a neutral mutation in a population is governed by the size of the population, the mutation rate, genetic drift and selective pressures on the alleles to which it is linked. It is understood that most of a species' genome, in the absence of selection, undergoes a steady accumulation of neutral mutations. If the environment later changes, a mutation that was originally neutral may become either deleterious or beneficial.
Individual genes can be affected by point mutations, also known as SNPs, in which a single base pair is altered. The substitution of a single base pair may or may not affect the function of the gene (see mutation). Deletions and insertions of a single or several base pairs usually results in a non-functional gene.[12] Note, however, that the insertion or deletion of three (or multiples of three) consecutive nucleotides may have little impact on the function of a gene, since the reading frame is maintained.
Mobile elements, transposons, make up a major fraction of the genomes of plants and animals and appear to have played a significant role in the evolution of genomes. These mobile insertional elements can jump within a genome and alter existing genes and gene networks to produce evolutionary change and diversity.
On the other hand, gene duplications, which may occur via a number of mechanisms, are believed to be one major source of raw material for evolving new genes as tens to hundreds of genes are duplicated in animal genomes every million years.[13] Most genes belong to larger "families" of genes derived from a common ancestral gene (two genes from a species that are in the same family are dubbed "paralogs"). Another mechanism causing gene duplication is intergenic recombination, particularly 'exon shuffling', i.e., an aberrant recombination that joins the 'upstream' part of one gene with the 'downstream' part of another. Genome duplications and chromosome duplications also appear to have served a significant role in evolution. Genome duplication has been the driving force in the Teleostei genome evolution, where up to four genome duplications are thought to have happened, resulting in species with more than 250 chromosomes.
Large chromosomal rearrangements do not necessarily change gene function, but do generally result in reproductive isolation, and, by definition, speciation (a species here is a group of organisms that can interbreed in nature, producing viable offspring as a result). An example of this mechanism occurred in our lineage, where two chromosomes fused to produce human chromosome 2. This fusion did not occur in the chimp, gorilla, or orangutan lineages, resulting in chimpanzees and the others retaining the primitive state where the two chromosomes are separate (chimps, gorillas, and orangutans have 24 pairs of chromosomes whereas humans have only 23 pairs).
Horizontal gene transfer and hybridization
Horizontal gene transfer (HGT) (or Lateral gene transfer) is any process in which an organism transfers genetic material (i.e. DNA) to another organism that is not its offspring. This mechanism allows for the transfer of genetic material between unrelated organisms of the same species or of different species and is a form of gene flow.

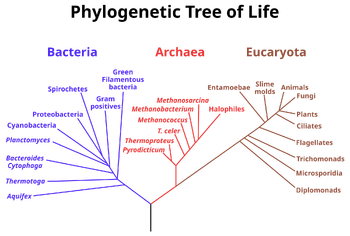
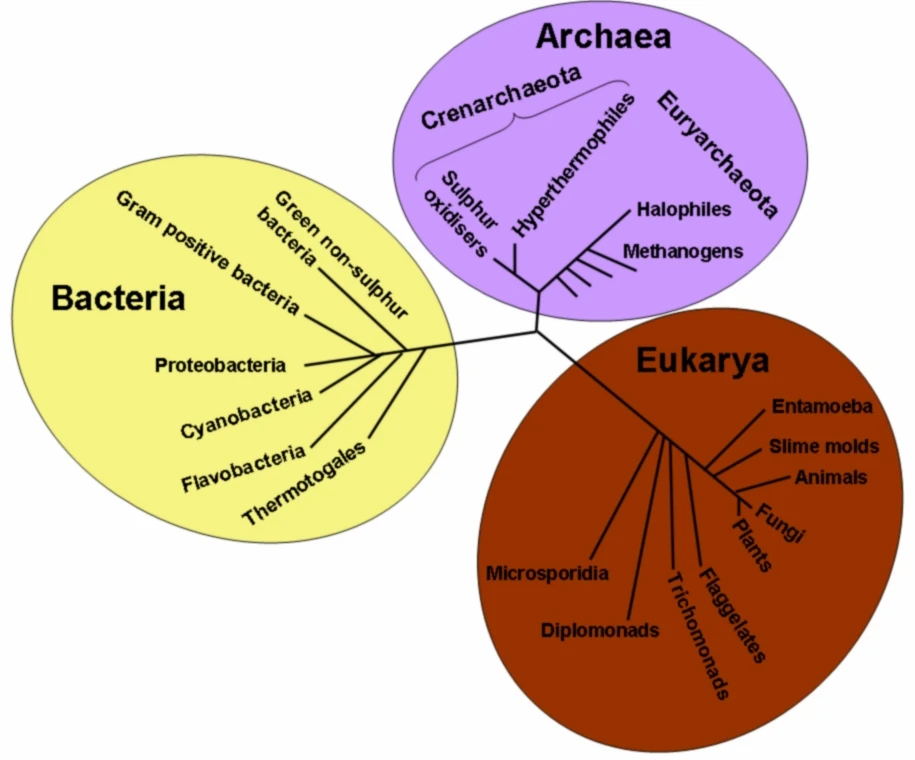
{kind=link}
A phylogenetic tree of all extant organisms, based on 16S rRNA gene sequence data, showing the evolutionary history of the three domains of life, bacteria, archaea and eukaryotes. Originally proposed by Carl Woese.
Many mechanisms for horizontal gene transfer have been observed, such as antigenic shift, reassortment, and hybridization. Viruses can transfer genes between species via transduction.[14] Bacteria can incorporate genes from other dead bacteria or plasmids via transformation, exchange genes with living bacteria via conjugation, and can have plasmids "set up residence separate from the host's genome".[15]
HGT has been shown to result in the spread of antibiotic resistance across bacterial populations.[16] Furthermore, findings indicate that HGT has been a major mechanism for prokaryotic and eukaryotic evolution.[17][18]
HGT complicates the inference of the phylogeny of life, as the original metaphor of a tree of life no longer fits. Rather, since genetic information is passed to other organisms and other species in addition to being passed from parent to offspring, "biologists [should] use the metaphor of a mosaic to describe the different histories combined in individual genomes and use [the] metaphor of a net to visualize the rich exchange and cooperative effects of HGT among microbes."[19]
Hybridization is highly significant in plant speciation, and one out of ten species of birds are known to hybridize. [20] There are also examples of hybridization in mammals and insects, however it often produces sterile offspring. Hybridization often produces variations that prove beneficial in extreme environments.
Cooperation
One of the most striking features of the natural world is the way in which genes, cells, and organisms cooperate to make higher order entities function. Generally mathematical models incorporating mutation and natural selection have been used to model adaptation and evolution. Recent trends now incorporate "game theory" as more applicable to generating reliable models [21]. This work and other studies have focused attention on cooperation as a fundamental property needed for evolution to construct new levels of organization. Selfish replicators sacrificing their own reproductive potential to cooperate seems paradoxical in a competitive world, however a number of mechanisms have demonstrated the capacity to generate cooperation: like kin selection, direct reciprocity, indirect reciprocity, network reciprocity, and group selection. The ubiquity of cooperation in the natural world and studies from the last twenty-five years[22] reveal cooperation as a significant principle in constructive evolution[23] and in 2006 it was recognised as the third fundamental principle in evolution alongside variation and selection[24].
- See also: Current Research in Evolutionary Biology and Reciprocity (evolution)
Mechanisms of evolution
Selection and adaptation
{kind=link}
A peacock's tail is the canonical example of sexual selection.
Natural selection refers to differences in survival and reproduction. Differential mortality is the survival rate of individuals to their reproductive age. Differential fertility is the total genetic contribution to the next generation. Note that, whereas mutations and genetic drift are random, natural selection is not, as it preferentially selects for or against different mutations based on differential fitnesses. For example, rolling dice is random, but always picking the higher number on two rolled dice is not random. The central role of natural selection in evolutionary theory has given rise to a strong connection between that field and the study of ecology.
Natural selection can be subdivided into two categories:
- Ecological selection occurs when organisms that survive and reproduce increase the frequency of their genes in the gene pool over those that do not survive.
- Sexual selection occurs when organisms which are more attractive to the opposite sex because of their features reproduce more and thus increase the frequency of those features in the gene pool.
Natural selection also operates on mutations in several different ways:
- Positive or directional selection increases the frequency of a beneficial mutation, or pushes the mean in either direction.
- Purifying or stabilizing selection maintains a common trait in the population by decreasing the frequency of harmful mutations and weeding them out of the population. "Living fossils" are arguably the product of stabilizing selection, as their form and traits have remained virtually identical over a long period. It is argued that stabilizing selection is the most common form of natural selection.
- Balancing selection maintains variation within a population through a number of mechanisms, including:
- Heterozygote advantage or overdominance, where the heterozygote is more fit than either of the homozygous forms (exemplified by human sickle cell trait conferring resistance to malaria)
- Frequency-dependent selection, where rare variants either have increased fitness or decreased fitness, because of their rarity.
- Disruptive selection favors both extremes, and results in a bimodal distribution of gene frequency. The mean may or may not shift.
- Selective sweeps describe the affect of selection acting on linked alleles. It comes in two forms:
- Background selection occurs when a deleterious mutation is selected against, and linked mutations are eliminated along with the deleterious variant, resulting in lower genetic polymorphism in the surrounding region.
- Genetic hitchhiking occurs when a beneficial allele is selected for, and linked alleles, which can be neutral or beneficial, are pushed towards fixation along with the beneficial allele.
- Artificial selection refers to the intentional or unintentional directed breeding of a species by human intervention. Many examples derive from selective breeding and domestication of animals and plants for agriculture, horticulture, livestock breeding, and pets. Such selection can induce dramatic changes in a breed in a relatively short period of time; however, because humans are only part of any species' environment, the degree to which any change can be ascribed to human intervention may be difficult to determine. Deliberate artificial selection within human populations is a controversial enterprise known as eugenics.
Through the process of natural selection, organisms become better adapted to their environments. Adaptation is any evolutionary process that increases the fitness of individuals, or sometimes the trait that confers increased fitness, e.g. a stronger prehensile tail or greater visual acuity. Note that adaptation is context-sensitive; a trait that increases fitness in one environment may decrease it in another.
Evolution does not act in a linear direction towards a pre-defined "goal" — it only responds to various types of adaptationary changes. The belief in a teleological evolution of this goal-directed sort is known as orthogenesis, and is not supported by the scientific understanding of evolution. Specific mutations do not arise because they would be beneficial: future needs do not drive the process of mutation (mutations are random with respect to fitness).
Most biologists believe that adaptation occurs through the accumulation (via natural selection) of many beneifical mutations of small effect. However, macromutation is a potential (rare) alternative process for (external) adaptation: here, a mutation occurs that has a relatively large-scale effect on the external phenotype that happens to be beneficial. This sudden change could leave the organism "uncoordinated" internally. Internal selection could then become a factor, improving internal coadaptation.
Recombination
In asexual organisms, alleles on the same chromosome will always be inherited together — they are linked, by virtue of being on the same DNA molecule. However, sexual organisms, in the production of gametes, shuffle linked alleles on homologous chromosomes inherited from the parents via meiotic recombination. This shuffling allows independent assortment of alleles (mutations) in genes to be propagated in the population independently. This allows bad mutations to be purged and beneficial mutations to be retained more efficiently than in asexual populations.
However, the meiotic recombination rate is not very high - on the order of one crossover (recombination event between homologous chromosomes) per chromosome arm per generation. Therefore, linked alleles are not perfectly shuffled away from each other, but tend to be inherited together. This tendency may be measured by comparing the co-occurrence of two alleles, usually quantified as linkage disequilibrium (LD). A set of alleles that are often co-propagated is called a haplotype. Strong haplotype blocks can be a product of strong positive selection.
Recombination is mildly mutagenic, which is one of the proposed reasons why it occurs with limited frequency. Recombination also breaks up gene combinations that have been successful in previous generations, and hence should be opposed by selection. However, recombination could be favoured by negative frequency-dependent selection (this is when rare variants increase in frequency) because it leads to more individuals with new and rare gene combinations being produced.
When alleles cannot be separated by recombination (for example in mammalian Y chromosomes), there is an observable reduction in effective population size, known as the Hill-Robertson effect, and the successive establishment of bad mutations, known as Muller's ratchet.
Genetic drift
Genetic drift, which is more pronounced the smaller the population is, is the change in allele frequency from one generation to the next due to sampling variance. The frequency of an allele in the offspring generation will vary according to a probability distribution of the frequency of the allele in the parent generation. Thus, over time even in the absence of selection upon the alleles, allele frequencies tend to "drift" upward or downward, eventually becoming lost or fixed - that is, going to 0% or 100% frequency. Thus, fluctuations in allele frequency between successive generations may result in some alleles disappearing from the population due to chance alone. Two separate populations that begin with the same allele frequencies therefore might drift apart by random fluctuation into two divergent populations with different allele sets (for example, alleles present in one population could be absent in the other, or vice versa).
Gene flow
- Main article: Gene flow
{kind=link}
Map of the world showing distribution of camelids. Solid black lines indicate possible migration routes.
Gene flow (also called gene admixture or simply migration) is the exchange of genetic variation between populations, when geography and culture are not obstacles. Ernst Mayr thought that gene flow is likely to be homogenising, and therefore counteract selective adaptation. Where there are obstacles to gene flow, the situation is termed reproductive isolation and is considered to be necessary for speciation.
The free movement of alleles through a population may also be impeded by population structure. For example, most real-world populations are not actually fully interbreeding; geographic proximity has a strong influence on the movement of alleles within the population.
Population structure
Population structure is the size and geographical distribution of a population. It has profound effects on possible mechanisms of evolution. The consequence of genetic drift depends strongly on the size of the population (generally abbreviated as N): drift is important in small mating populations (see Founder effect and Population bottleneck), where chance fluctuations from generation to generation can be large. The relative importance of natural selection and genetic drift in determining the fate of new mutations also depends on the population size and the strength of selection, s: when N times s is small, genetic drift predominates. When N times s is large, selection predominates. Thus, natural selection is predominant in large populations, while genetic drift is in small populations. Finally, the time for an allele to become fixed in the population by genetic drift (that is, for all individuals in the population to carry that allele) depends on population size -- smaller populations require a shorter time for fixation.
An example of the effect of population structure is the founder effect, resulting from a migration or population bottleneck, in which a population temporarily has very few individuals, and therefore loses much genetic variation. In this case, a single, rare allele may suddenly increase very rapidly in frequency within a specific population if it happened to be prevalent in a small number of "founder" individuals. The frequency of the allele in the resulting population can be much higher than otherwise expected, especially for deleterious, disease-causing alleles (because competition between members of the population is minimal, reducing by some degree the importance of fitness). Since population size has a profound effect on the relative strengths of genetic drift and natural selection, changes in population size can alter the dynamics of these processes considerably.
Speciation and extinction


{kind=link}
An Allosaurus skeleton.
Speciation is the process by which new biological species arise. Evolutionary biologist and a father of the modern synthesis, Ernst Mayr, defined species as "groups of actually or potentially interbreeding natural populations, which are reproductively isolated from other such groups." [25] This may take place by various mechanisms. Allopatric speciation occurs in populations that become isolated geographically, such as by habitat fragmentation or migration.[26] Sympatric speciation occurs when new species emerge in the same geographic area.[27][28] Ernst Mayr's peripatric speciation is a type of speciation that exists in between the extremes of allopatry and sympatry. Peripatric speciation is a critical underpinning of the theory of punctuated equilibrium. An example of rapid sympatric speciation can be eloquently represented in the triangle of U; where new species of Brassica sp. have been made by the fusing of separate genomes from related plants.
Extinction is the disappearance of species (i.e. gene pools). The moment of extinction generally occurs at the death of the last individual of that species. Extinction is not an unusual event in geological time — species are created by speciation, and disappear through extinction. The Permian-Triassic extinction event was the Earth's most severe extinction event, rendering extinct 90% of all marine species and 70% of terrestrial vertebrate species. In the Cretaceous-Tertiary extinction event many forms of life perished (including approximately 50% of all genera), the most often mentioned among them being the extinction of the (non-avian) dinosaurs.
Evidence of evolution
{kind=link}
Tiktaalik in context: one of many species that track the evolutionary development of fish fins into tetrapod limbs.
Evolution has left numerous records that reveal the history of different species. Fossils, together with the comparative anatomy of present-day plants and animals, constitute the morphological, or anatomical, record. By comparing the anatomies of both modern and extinct species, paleontologists can infer the lineages of those species. Important fossil evidence includes the connection of distinct classes of organisms by so-called "transitional" species, such as the Archaeopteryx, which provided early evidence for intermediate species between dinosaurs and birds,[29] and the recently-discovered Tiktaalik, which clarifies the development from fish to terrestrial animals with four limbs.[30]
The development of molecular genetics, and particularly of DNA sequencing, has allowed biologists to study the record of evolution left in organisms' genetic structures. The degrees of similarity and difference in the DNA sequences of modern species allows geneticists to reconstruct their lineages. It is from DNA sequence comparisons that figures such as the 95% genotypic similarity between humans and chimpanzees are obtained.[31][32]
Additional evidence of ancestry includes idiosyncratic structures present in certain organisms, such as the panda's "thumb", which indicate how an organism's evolutionary lineage constrains its adaptive development. In addition, vestigial structures such as the vestigial limbs on pythons; the ball-and-socket joints for "ghost" femurs in manatees; the degenerate eyes of blind cave-dwelling fish; and our wisdom teeth, coccyx, and vermiform appendix; are also evidences of descent with modification.
Developmental biology also provides evidence for evolution and common descent. For example, although tails are useless to humans, our embryos - just like those of pigs, dogs, cats, mice, and so on - have a postanal tail. In addition, humans have only a single aortic arch, but our embryos start off with the same arrangement as embryonic fishes, where there are six pairs of aortic arches: during further embryonic development in humans, much remodeling occurs to convert the fish arrangement of aortic arches into the human arrangement.
Other evidence used to demonstrate evolutionary lineages includes the geographical distribution of species (biogeography). For instance, monotremes, such as platypus, and most marsupials, like kangaroos or koalas, are found only in Australia showing that their common ancestor with placental mammals lived before the submerging of the ancient land bridge between Australia and Asia.
Scientists correlate all of the above evidence, drawn from paleontology, anatomy, genetics, development/embryology, and biogeography, with other information about the history of Earth. For instance, paleoclimatology attests to periodic ice ages during which the world's climate was much cooler, and these are often found to match up with the spread of species which are better-equipped to deal with the cold, such as the woolly mammoth.
Morphological evidence
{kind=link}
Letter c in the picture indicates the undeveloped hind legs of a baleen whale, vestigial remnants of its terrestrial ancestors.
Fossils are critical evidence for estimating when various lineages originated. Since fossilization of an organism is an uncommon occurrence, usually requiring hard parts (like teeth, bone or pollen), the fossil record is traditionally thought to provide only sparse and intermittent information about ancestral lineages. Fossilization of organisms without hard body parts is rare, but happens under unusual circumstances, such as rapid burial, low oxygen environments, or microbial action.[33]
The fossil record provides several types of data important to the study of evolution. First, the fossil record contains the earliest known examples of life itself, as well as the earliest occurrences of individual lineages. For example, the first complex, macroscopic animals date from the Early Cambrian period, approximately 530 million years ago. Second, the records of individual species yield information regarding the patterns and rates of evolution, showing for example whether certain species evolve into new species gradually and incrementally (phyletic gradualism), or in relatively brief intervals of geologic time after long spans of stasus(punctuated equilibria). Thirdly, the fossil record is a document of large scale patterns and events in the history of life, many of which have influenced the evolutionary history of numerous lineages. For example, mass extinctions frequently resulted in the loss of entire groups of species, such as the (non-avian) dinosaurs, while leaving others relatively unscathed. Recently, molecular biologists have used the time since divergence of related lineages to calibrate the rate at which mutations accumulate, and at which the genomes of different lineages evolve.
Phylogenies indicate the ancestry of species, and are based on evidences from the fossil record, comparative molecular genetics, comparative anatomy, and comparative embryology. Phylogenetic trees constructed from such information help support hypotheses about whether certain structures shared by two (or more) species are homologous (in which case they were inherited from a common ancestor) or are homoplasious (in which case they arose independently). A commonly used example of homologous structures are Vertebrate limbs, which have very similar skeletal structures (suggesting common ancestry) but perform different functions. For example, a human hand is used for grasping, a bat's wing is used for flying, a dolphin's fin is used in swimming, and a frog's front legs are used for locomotion: yet they all share the same general internal, skeletal structure. All of the evidence points to vertebrate limbs being homologous, and thus such homologous structures also support evolution and common descent.
Molecular evidence
Comparison of the DNA sequences allows organisms to be grouped by sequence similarity, and the resulting phylogenetic trees are typically congruent with traditional taxonomy, and are often used to strengthen or correct taxonomic classifications. Sequence comparison is considered a measure robust enough to be used to correct erroneous assumptions in the phylogenetic tree in instances where other evidence is scarce. For example, neutral human DNA sequences are approximately 1.2% divergent (based on substitutions) from those of their nearest genetic relative, the chimpanzee, 1.6% from gorillas, and 6.6% from baboons.[34] Genetic sequence evidence thus allows inference and quantification of genetic relatedness between humans and other apes.[35][36] The sequence of the 16S rRNA gene, a vital gene encoding a part of the ribosome, was used to find the broad phylogenetic relationships between all extant life. The analysis, originally done by Carl Woese, resulted in the three-domain system, arguing for two major splits in the early evolution of life. The first split led to modern Bacteria and the subsequent split led to modern Archaea and Eukarya.
The proteomic evidence also supports the universal ancestry of life. Vital proteins, such as those in the ribosome, as well as DNA polymerase, and RNA polymerase, are found in the most primitive bacteria to the most complex mammals. The core part of the protein is conserved across all lineages of life, serving similar functions. Higher organisms have evolved additional protein subunits, largely affecting the regulation and protein-protein interaction of the core. Other overarching similarities between all lineages of extant organisms, such as DNA, RNA, amino acids, and the lipid bilayer, give support to the theory of common descent. The chirality of DNA, RNA, and amino acids is conserved across all known life. As there is no functional advantage to right or left handed molecular chirality, the simplest hypothesis is that the choice was made randomly in the early beginnings of life and passed on to all extant life through common descent.
There is also a large body of molecular evidence for a number of different mechanisms for large evolutionary changes, among them genome and gene duplication, horizontal gene transfer, recombination, and endosymbiosis. These mechanisms have been, or are being incorporated into the Modern Evolutionary Synthesis :
- Gene and genome duplication facilitates rapid evolution by providing substantial quantities of genetic material under weak or no selective constraints.
- Horizontal gene transfer, the process in which an organism transfers genetic material (i.e. DNA) to another cell that is not its offspring, allows for large sudden evolutionary leaps in a species by incorporating beneficial genes evolved in another species.
- Recombination is both capable of re-assorting large numbers of different alleles, and establishing reproductive isolation.
- The Endosymbiotic theory explains the origin of mitochondria and plastids (e.g. chloroplasts), which are organelles of eukaryotic cells, as the incorporation of an ancient prokaryotic cell into an ancient proto-eukaryotic cell. Rather than evolving eukaryotic organelles slowly, this scientific theory offers a mechanism for large evolutionary changes by incorporating the genetic material and biochemical composition of a separate species. This evolutionary mechanism has been observed. Hatena, a protist, is an extant organism that is undergoing endosymbiotic evolution.[37][38]
Further evidence for reconstructing ancestral lineages comes from so-called junk DNA such as pseudogenes, i.e., 'dead' genes, which steadily accumulate mutations.[39]. Because humans and chimps share several pseudogenes, and in some cases also share the very same mutations that make the genes non-functional, our descent from a ancestor we share with chimps is further supported (the best explanation for multiple shared defects is common ancestry).
Since metabolic processes do not leave fossils, research into the evolution of the basic cellular processes is done largely by comparison of existing organisms' processes. Many lineages diverged when new metabolic processes appeared, and it is theoretically possible to determine when certain metabolic processes appeared by comparing the traits of the descendants of a common ancestor or by detecting their physical manifestations. As an example, the appearance of oxygen in the earth's atmosphere is linked to the evolution of photosynthesis.
Theoretical evidence
Mathematical models of evolution, pioneered by the likes of Sewall Wright, Ronald Fisher and J. B. S. Haldane, and extended via diffusion theory by Motoo Kimura, allow predictions about the genetic structure of evolving populations to be made. Direct examination of the genetic structure of modern populations via DNA sequencing has recently allowed verification of many of these predictions. For example, the Out of Africa theory of human origins, which states that modern humans developed in Africa and a small sub-population migrated out (undergoing a population bottleneck), implies that modern populations should show the signatures of this migration pattern. Specifically, post-bottleneck populations (Europeans and Asians) should show lower overall genetic diversity and a more uniform distribution of allele frequencies compared to the African population.[40] Both of these predictions are borne out by actual data from a number of studies. [41][42]
Ancestry of organisms
- See also: Common descent
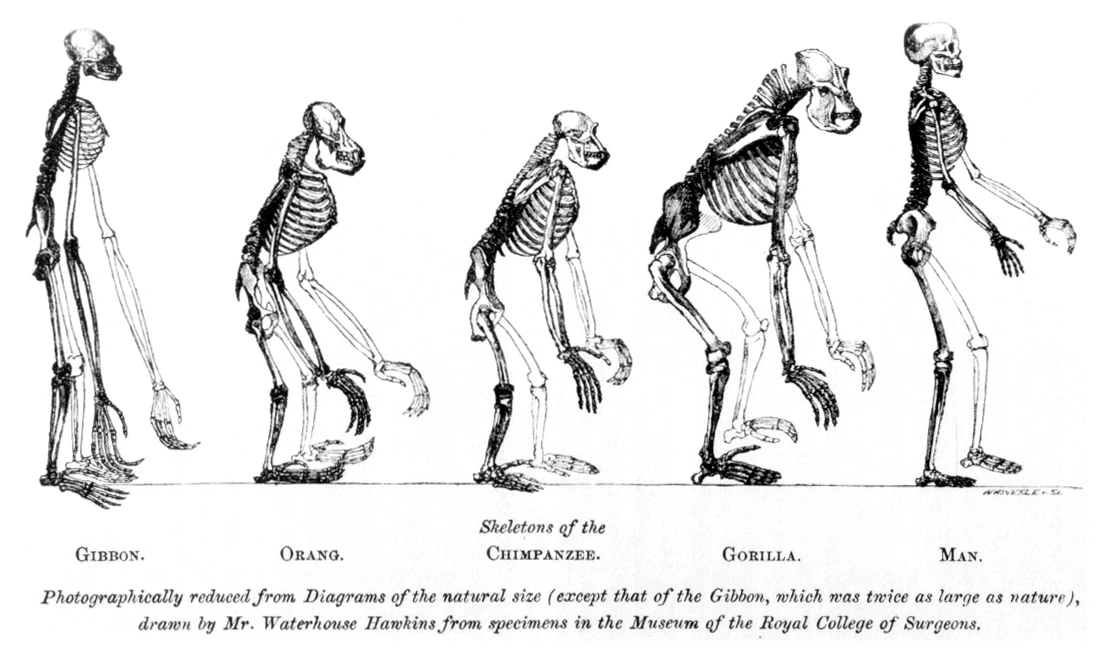

{kind=link}
Morphologic similarities in the Hominidae family is evidence of common descent.
In biology, the theory of universal common descent proposes that all organisms on Earth are descended from a common ancestor or ancestral gene pool.
Evidence for common descent is inferred from traits shared between all living organisms. In Darwin's day, the evidence of shared traits was based solely on visible observation of morphologic similarities, such as the fact that all birds, even those which do not fly, have wings. Today, there is strong evidence from molecular biology that all organisms have a common ancestor. For example, every living cell makes use of nucleic acids as its genetic material, and uses the same set of 20 common amino acids as the building blocks for proteins. All organisms also use the same genetic code (with some extremely rare and minor deviations) to translate nucleic acid sequences into proteins. Numerous other universals exist, such as all cells using ribosomes composed of one large and one small subunit, with the first amino acid brought to the ribosome during translation being methionine (or formyl-methionine). These core cellular process are so universally conserved that a human gene can be transferred into bacteria and they will produce the correct, functional, human protein. The universality of these traits strongly suggests common ancestry, because the selection of many of these traits seems arbitrary: for example, there appears to be no inherent need for all cells to use methionine, instead of any of the other 19 common amino acids, to initiate protein synthesis.
Information about the early development of life includes input from the fields of geology and planetary science. These sciences provide information about the history of Earth and the changes produced by life. However, a great deal of information about the early Earth has been destroyed by geological processes over the course of time.
History of life
The chemical evolution (or abiogenesis) from self-catalytic chemical reactions to life (see Origin of life) is not a part of biological evolution, but it is unclear at which point such increasingly complex sets of reactions became what we would consider, today, to be living organisms.
Not much is yet known about the earliest developments in life. However, it is unmistakably clear that all existing organisms share certain traits, including cellular structure and the same genetic code. Most scientists interpret this to mean all existing organisms share a common ancestor, which had already developed the most fundamental cellular processes, but there is no scientific consensus about the relationship of the three domains of life (Archaea, Bacteria, Eukarya) or the origin of life. Attempts to shed light on the earliest history of life generally focus on the behavior of macromolecules, particularly RNA, and the behavior of complex systems.


{kind=link}
Precambrian stromatolites in the Siyeh Formation, Glacier National Park. In 2002, William Schopf of UCLA published a controversial paper in the journal Nature arguing that formations such as this possess 3.5 billion year old fossilized algae microbes. If true, they would be the earliest known life on earth.
The emergence of oxygenic photosynthesis (around 3 billion years ago) and the subsequent emergence of an oxygen-rich, non-reducing atmosphere can be traced through the formation of banded iron deposits, and later red beds of iron oxides. This was a necessary prerequisite for the development of aerobic cellular respiration, believed to have emerged around 2 billion years ago.
In the last billion years, simple multicellular plants and animals began to appear in the oceans. Soon after the emergence of the first animals, the Cambrian explosion (a period of unrivaled and remarkable, but brief, organismal diversity documented in the fossils found at the Burgess Shale) saw the creation of all the major body plans, or phyla, of modern animals. This event is now believed to have been triggered by the development of the Hox genes. About 500 million years ago, plants and fungi colonized the land, and were soon followed by arthropods and other animals, leading to the development of land ecosystems with which we are familiar.
The evolutionary process may be exceedingly slow. Fossil evidence indicates that the diversity and complexity of modern life has developed over much of the history of Earth. Geological evidence indicates that Earth is approximately 4.57 billion years old. However, studies on guppies by David Reznick at the University of California, Riverside, have shown that the rate of evolution through natural selection can proceed 10 thousand to 10 million times faster than what is indicated in the fossil record.[43] But such comparative studies are invariably biased by disparities in the time scales over which evolutionary change is measured in the laboratory, field experiments, and the fossil record.
The ancestry of living organisms has traditionally been reconstructed from morphology, but is increasingly supplemented with phylogenetics — the reconstruction of phylogenies by the comparison of genetic (usually DNA) sequence.[44] Biologist Gogarten suggests that "the original metaphor of a tree no longer fits the data from recent genome research", and that therefore "biologists [should] use the metaphor of a mosaic to describe the different histories combined in individual genomes and use [the] metaphor of a net to visualize the rich exchange and cooperative effects of HGT among microbes".[45]
History of evolutionary thought
{kind=link}
Charles Darwin in 1854, five years before publishing The Origin of Species.
The idea of biological evolution has existed since ancient times, notably among Greek philosophers such as Anaximander and Epicurus and Indian philosophers such as Patañjali. However, scientific theories of evolution were not proposed until the 18th and 19th centuries, by scientists such as Jean-Baptiste Lamarck and Charles Darwin.
The transmutation of species was accepted by many scientists before 1859, but Charles Darwin's On the Origin of Species by Means of Natural Selection provided the first convincing exposition[46] of a mechanism by which evolutionary change could occur: natural selection. After many years of working in private on his theory, Darwin was motivated to publish his work on evolution when he received a letter from Alfred Russel Wallace in which Wallace revealed his own, independent discovery of natural selection. Accordingly, Wallace is given shared credit for originating the theory.[47][48]
The publication of Darwin's book sparked a great deal of scientific and social debate. Although the occurrence of biological evolution of some sort came to be widely accepted by scientists, Darwin's specific ideas about evolution — that it occurred gradually, through natural selection — were actively attacked and contested. Additionally, while Darwin was able to observe variation, and to infer natural selection and thereby adaptation, he was unable to explain how variation might arise or be altered over generations.[49]

{kind=link}
Gregor Mendel's work on the inheritance of traits in pea plants laid the foundation for genetics.
Work on plant hybridity by a contemporary of Darwin's, Gregor Mendel, revealed that certain traits in peas occurred in discrete forms (that is, they were either one distinct trait or another, such as "round" or "wrinkled") and were inherited in a well-defined and predictable manner.[50] When Mendel's work was "rediscovered" in 1901, it was initially interpreted as supporting an anti-Darwinian "jumping", saltationist form of evolution, and contradicting the biometricians' gradualism.[51]
The simple version of the theory of early Mendelians soon became incorporated into the classical genetics of Thomas Hunt Morgan and his school, which thoroughly grounded and articulated the applications of Mendelian laws to biology. Eventually, it was shown that a rigorous statistical approach to Mendelism was reconcilable with the data of the biometricians by the work of statistician and population geneticist R.A. Fisher in the 1930s. Following this, the work of population geneticists and zoologists in the 1930s and 1940s synthesized Darwinian evolution with genetics, creating the modern evolutionary synthesis.[50] Genes were then still theoretical entities, and many paleontologists and embryologists were inclined to dismiss them as being of no, or minor, importance, but subsequent advancements have made genetics a key aspect of evolutionary biology.[52]
{kind=link}
Stephen Jay Gould was a major proponent of punctuated equilibrium.
The most significant recent developments in evolutionary biology have been the improved understanding of and advances in genetics.[53] In the 1940s, following up on Griffith's experiment, Avery, MacLeod and McCarty definitively identified DNA (deoxyribonucleic acid) as the "transforming principle" responsible for transmitting genetic information. In 1953, Francis Crick and James D. Watson published their famous paper on the structure of DNA, based on the research of Rosalind Franklin and Maurice Wilkins. These developments ignited the era of molecular biology and transformed the understanding of evolution into a molecular process (see molecular evolution): the mutation of segments of DNA. George C. Williams's 1966 Adaptation and natural selection: A Critique of some Current Evolutionary Thought and Richard Dawkins' The Selfish Gene marked a departure from the idea of groups or organisms as units of selection toward the modern gene-centered view of evolution. In the mid-1970s, Motoo Kimura formulated the neutral theory of molecular evolution, a significant departure from the consensus view of evolution, as it considered genetic drift, rather than natural selection to be the predominant mode of evolution.
Debates over various aspects of how evolution occurs have continued. Two prominent debates are over the theory of punctuated equilibrium, proposed in 1972 by paleontologists Niles Eldredge and Stephen Jay Gould to explain the paucity of gradual transitions between species in the fossil record, as well as the absence of change or stasis that is observed over significant intervals of time; and also the Neutralist-Selectionist debate.
Modern synthesis
Charles Darwin was able to observe variation, infer natural selection and thereby adaptation, but did not know the basis of heritability. He could not explain how organisms might change over generations. It also seemed that when two individuals were crossed, their traits must be blended in the progeny, so that eventually all variation would be lost.
The blending problem was solved when the population geneticists R.A. Fisher, Sewall Wright, and J. B. S. Haldane, married Darwinian evolutionary theory to population genetics, which was based on Mendelian genetics (genes as discrete units of heredity).
The problem of what the mechanisms might be was solved in principle with the identification of DNA as the genetic material by Oswald Avery and colleagues, and the articulation of the double-helical structure of DNA by James Watson and Francis Crick provided a physical basis for the notion that genes were encoded in DNA.
Study of evolution
Academic disciplines
Scholars in a number of academic disciplines continue to document examples of evolution, contributing to a deeper understanding of its underlying mechanisms. Every subdiscipline within biology both informs and is informed by knowledge of the details of evolution, such as in ecological genetics, human evolution, molecular evolution, and phylogenetics. Areas of mathematics (such as bioinformatics), physics, chemistry, and other fields all make important contributions to current understanding of evolutionary mechanisms. Even disciplines as far removed as geology and sociology play a part, since the process of biological evolution has coincided in time and space with the development of both the Earth and human civilization.
Evolutionary biology is a subdiscipline of biology concerned with the origin and descent of species, as well as their changes over time. It was originally an interdisciplinary field including scientists from many traditional taxonomically-oriented disciplines. For example, it generally includes scientists who may have a specialist training in particular organisms, such as mammalogy, ornithology, or herpetology, but who use those organisms to answer general questions in evolution. Evolutionary biology as an academic discipline in its own right emerged as a result of the modern evolutionary synthesis in the 1930s and 1940s. It was not until the 1970s and 1980s, however, that a significant number of universities had departments that specifically included the term evolutionary biology in their titles.
Evolutionary developmental biology (informally, evo-devo) is a field of biology that compares the developmental processes of different animals in an attempt to determine the ancestral relationship between organisms and how developmental processes evolved. The discovery of genes regulating development in model organisms allowed for comparisons to be made with genes and genetic networks of related organisms.
Physical anthropology emerged in the late 19th century as the study of human osteology, and the fossilized skeletal remains of other hominids. At that time, anthropologists debated whether their evidence supported Darwin's claims, because skeletal remains revealed temporal and spatial variation among hominids, but Darwin had not offered an explanation of the specific mechanisms that produce variation. With the recognition of Mendelian genetics and the rise of the modern synthesis, however, evolution became both the fundamental conceptual framework for, and the object of study of, physical anthropologists. In addition to studying skeletal remains, they began to study genetic variation among human populations (population genetics); thus, some physical anthropologists began calling themselves biological anthropologists.
Misunderstandings about modern evolutionary biology
Although the modern synthesis is a major achievement of modern science, some aspects of it are often misunderstood. These misunderstandings have hindered acceptance of the modern synthesis,[54][55] most notably in the United States.[56] Some of the most common misunderstandings are outlined in this section.
Distinctions between theory and fact
- Further information: Scientific Theory
- See also: Theory vs. Fact
Some critics of evolution claim that it is merely a theory. For example, Ronald Reagan quiped during the 1980 presidential campaign, "Well, [evolution] is a theory--it is a scientific theory only, and it has in recent years been challenged in the world of science and is not yet believed in the scientific community to be as infallible as it was once believed." Cite error: Invalid <ref> tag; invalid names, e.g. too manyCite error: Invalid <ref> tag; invalid names, e.g. too many This criticism makes two claims: that evolution is a theory and therefore not a fact, and that theories are less established than facts (and less well supported than other beliefs). In other words, laypeople often use the word "theory" to signify "conjecture", "speculation", or "opinion". In science however, a theory is a model of the world (or some portion of it) that makes predictions that can be tested through controlled experiments.
A second linguistic confusion has arisen around the word "fact". Fact is often used by scientists to refer to empirical data, objective verifiable observations. But fact is also used in a wider sense to include any hypothesis for which there is overwhelming evidence. In this usage "the sun is at the center of the solar system" and "objects fall due to gravity" are considered to be facts.
Moreover the word evolution may refer to the observed fact, or it may refer to a theory which explains it. When "evolution" is used to describe a fact, it refers to the observations that populations of one species of organism do, over time, change into new, or several new, species. In this sense, evolution occurs whenever a new strain of bacterium evolves that is resistant to antibodies that had been lethal to prior strains. Another clear case of evolution as fact involves the hawthorn fly.
When "evolution" is used to describe a theory, it refers to an explanation for why and a model of how evolution occurs. An example of evolution as theory is the modern synthesis of Darwin and Wallace's theory of natural selection and Mendel's principles of genetics. As with any scientific theory, the modern synthesis is constantly debated, tested, and refined by scientists. There is an overwhelming consensus in the scientific community that it remains the only robust model that accounts for the known facts concerning evolution.
| Gravity | Evolution |
|---|---|
| Things falling is an observation of the pull of bodies towards each other. | Fruit flies changing generation to generation is an observation of generational organism change. |
| Bodies pulling towards each other is called gravity. | Organisms changing generation to generation is called evolution. |
| Gravity is a "fact". | Evolution is a "fact". |
| The "facts" of gravity require an explanation. | The "facts" of evolution require an explanation. |
| Aristotle and Galileo created explanations of the "fact" of gravity. These are now obsolete explanations. Newton created an explanation which is substantially correct as far as it goes but turned out to require refinement. | Lamarckism, Transmutationism and Orthogenesis were created as explanations of the "fact" of evolution. These are now obsolete explanations. Darwin created an explanation which is substantially correct as far as it goes, but turned out to require refinement. |
| Einstein's explanation is a refinement of Newton's explanation of gravity. Einstein's explanation is currently the most accepted explanation of the "fact" of gravity. | The Neo-Darwinist explanation is a refinement of Darwin's explanation of evolution. Neo-Darwinism is currently the most accepted explanation of the "fact" of evolution. |
| Einstein's explanation of the "fact" of gravity is called the Theory of Gravity. | The Neo-Darwinist explanation of the "fact" of evolution is called the Theory of Evolution. |
| Gravity is a "fact" and a "theory." | Evolution is a "fact" and a "theory." |
Evolution, complexity, and devolution
One of the most common misunderstandings about evolution is that one species can be "more highly evolved" than another; that evolution is necessarily progressive and/or leads to greater "complexity", or that its converse is "devolution."[57] Evolution is a non-directional process in the sense that it does not proceed toward any ultimate goal. Rather, evolution acts to optimize organisms to the conditions of their environment, whatever they might be, through the process of natural selection. Thus, evolution provides no assurance that later generations will be more complex than earlier generations, a fact that applies equally to human complexity and intelligence as it does to all other organisms on Earth today and in the past. The claim that evolution results in directional progress is not part of modern evolutionary theory; it derives from earlier belief systems which were held around the time when Darwin formulated his ideas.
In many cases evolution does proceed in the direction of increasing complexity.[58] The earliest organisms were maximally simple life forms. Evolution caused life to become more complex, since becoming simpler wasn't advantageous; indeed it was likely impossible since organisms of any lower complexity would be too simple to carry out basic life functions. Virtually the only avenues open for evolution in the earliest epoch of life’s history were toward more complex and specialized forms. Since then, evolved complexity is a pattern that has occurred again and again in numerous lineages, since it produces more specialized forms that are capable of exploiting environmental niches more effectively than competitors. Indeed, specialization through increased complexity allows an organism to define its own niche and evolve to it, thus also increasing biodiversity.
But complexity is not an ultimate goal of evolutionary processes, and the evidence supporting this abounds. Bacterial forms have remained comparatively simple over 3.8 billion years of evolution even though numerous forms have evolved and specialized to a variety of environments using diverse metabolic pathways. In these cases, complexity was not selected for even though substantial evolutionary change occurred. Simple organisms that rapidly metabolize energy-rich nutrients available in the environment are optimal forms that dominate the biosphere and the cycling of elements in it. Among eukarya much greater levels of complexity have evolved, but these organisms represent the smaller portion of the biomass on earth. Thus, evolution toward complexity is the exception and not the norm.
There are numerous examples of organisms evolving towards simpler forms. Vestigial structures constitute one line of evidence of lessening complexity evolving among eukarya. Pythons have a vestigial pelvis that is detached from the vertebra, and essentially floats in the abdominal cavity. This structure is functionless, and it degenerated from a functioning pelvis in a previous ancestor with legs. The Mexican Tetra (Astyanax mexicanus) has evolved from a previous sighted ancestor into a blind form. While the animal retains a rudimentary eye, the nerves and retina are degenerate and functionless.[59] The human appendix is a degenerate form of the caecum found in a wide variety of herbivorous vertebrates. In these animals, the caecum is a significant organ that functions to digest cellulosic plant material. In humans and hominoid apes, changes in diet have favored its degeneration. In each instance, natural selection has acted to produce decreasing complexity over time because the maintenance of useless organs represents a drain on the organism’s energy resources and they are thus subjected to negative selective pressure.[60]
Similarly, horses evolved from morphologically more complex ancestors with five toes on their feet[61]. Over time, the fossil record shows a progression to reduced complexity that has produced the single toe (hoof) observed today. In the case of this animal, a simpler design is superior for the conditions under which it must survive. Many parasitic and symbiotic species have evolved to simpler forms from more complex forms to the point that they are incapable of existing outside the bodies of their hosts or symbionts. In these cases, reduced complexity offers a selective advantage but comes with the cost of surrendering independence.
There is no guarantee that any particular organism existing today will become more complex in the future. In fact, natural selection will only favor increasing complexity if it increases an organism’s chance of survival and ability to produce viable offspring that live long enough to reach sexual maturity. The same mechanism will also favor lower complexity in traits if that confers a selective advantage in the organism's environment.[62]
Speciation
{kind=link}
The existence of several different, but related, finches on the Galápagos Islands is evidence of the occurrence of speciation.
It is sometimes claimed that speciation — the origin of new species — has never been directly observed, and thus evolution cannot be called sound science.[63] This is a misunderstanding of both science and evolution. First, scientific discovery does not occur solely through reproducible experiments; the principle of uniformitarianism allows natural scientists to infer causes through their empirical effects. Moreover, since the publication of On the Origin of Species scientists have confirmed Darwin's hypothesis by data gathered from sources that did not exist in his day, such as DNA similarity among species and new fossil discoveries. Finally, speciation has actually been directly observed.[64] (See the hawthorn fly example.) Further, there are a number of examples of speciation in plants,[65] and differences in ectodysplasin alleles in stickleback fish speciation has developed as a supermodel for studying gene alterations and speciation.[66]
A variation of this assertion, that microevolution has been directly observed and macroevolution has not, is subject to the same counterarguments. However, it is generally accepted that macroevolution uses the same mechanisms of change as those already observed in microevolution.
Entropy and life
It is claimed that evolution, by increasing complexity, violates the second law of thermodynamics. This law posits that in an idealised isolated system, entropy will tend to increase or stay the same. Entropy is a measure of the dispersal of energy in a physical system so that it is not available to do mechanical work.[67] The claim ignores the fact that biological systems are not isolated systems. Life inherently involves open systems, not isolated systems, as all organisms exchange energy and matter with their environment, and similarly the Earth receives energy from the Sun and emits energy back into space. Simple calculations[68] show that the Sun-Earth-space system does not violate the second law because the enormous increase in entropy due to the Sun and Earth radiating into space dwarfs the small decrease in entropy caused by the evolution of life.
Information
Some assert that evolution cannot create information, or that information can only be created by an intelligence. Physical information exists regardless of the presence of an intelligence, and evolution allows for new information whenever a novel mutation or gene duplication occurs and is kept. It does not need to be beneficial or visually apparent to be "information." However, even if those were requirements they would be satisfied with the appearance of nylon eating bacteria,[69] which required new enzymes to efficiently digest a material that never existed until the modern age.[70]
Japanese researchers demonstrated that nylon degrading ability can be obtained de novo in laboratory cultures of Pseudomonas aeruginosa strain POA, which initially had no enzymes capable of degrading nylon oligomers. This indicates that the ability of bacteria to digest nylon can evolve if proper artificial selection is applied.[71] Recently, the same group found X-ray crystal structure of the newly evolved nylon-digesting enzyme.[72] Using the structural results, the authors propose "that the amino acid replacements in the catalytic cleft of a preexisting esterase resulted in the evolution of the" nylon-digesting enzyme. This hypothesis still needs to be confirmed by detailed genetic studies.
Social and religious controversies


{kind=link}
A satirical 1871 image of Charles Darwin as a quadrupedal ape reflects part of the social controversy over whether humans and other apes share a common lineage.
Ever since the publication of The Origin of Species in 1859, the modern science of evolution has been a source of nearly constant controversy. In general, controversy has centered on the philosophical, cosmological, social, and religious implications of evolution, not on the science of evolution itself. The proposition that biological evolution occurs through the mechanism of natural selection has been almost completely uncontested within the scientific community for much of the 20th century.[73]
As Darwin recognized early on, perhaps the most controversial aspect of evolutionary thought is its applicability to human beings. Darwin attempted to show that the human race and anthropoid apes originally sprang from long extinct apelike ancestors in his 1871 book The Descent of Man. [74] The controversy was fueled in part by one of Darwin's most vigorous defenders, Thomas Huxley, who opined that Christianity is "a compound of some of the best and some of the worst elements of Paganism and Judaism, moulded in practice by the innate character of certain people of the Western World."[74][75]Cite error: Invalid <ref> tag; invalid names, e.g. too many The idea that all diversity in life, including human beings, arose through natural processes without a need for supernatural intervention poses difficulties for the belief in purpose inherent in most religious faiths — and especially for the Abrahamic religions. Many religious people are able to reconcile the science of evolution with their faith, or see no real conflict;[76] Judaism, Catholicism and Anglicanism are notable as major faith traditions whose adherents generally see no conflict between evolutionary theory and religious belief.[77][78] The idea that faith and evolution are compatible has been called theistic evolution. Another group of religious people, generally referred to as creationists, consider evolutionary origin beliefs to be incompatible with their faith, their religious texts and their perception of design in nature, and so cannot accept what they call "unguided evolution".
One particularly contentious topic evoked by evolution is the biological status of humanity. Whereas the classical religious view can broadly be characterized as a belief in the great chain of being (in which people are "above" the animals but slightly "below" the angels), the science of evolution shows that humans, as animals, share common ancestry with all other animals and are particularly close to primates such as chimpanzees, gibbons, gorillas, and orangutans. Some people find this offensive because it "degrades" humankind. A related conflict arises when critics combine the religious view of people's superior status with the mistaken notion that evolution is necessarily "progressive". If human beings are superior to animals yet evolved from them, these critics claim, inferior animals would not still exist. In fact, humans and other species share common ancestors, and it is these common ancestors that are extinct.[79]
In some countries — notably the United States — these and other tensions between religion and science have fueled what has been called the creation-evolution controversy, which, among other things, has generated struggles over teaching curricula. While many other fields of science, such as cosmology[80] and earth science[81] also conflict with a literal interpretation of many religious texts, evolutionary studies in biology have borne the brunt of these debates. It also affects the public school system, where religion is assumed to have no effect on the teachings.
Evolution has been used to support philosophical and ethical choices which most contemporary scientists consider were neither mandated by evolution nor supported by science. For example, the eugenic ideas of Francis Galton were developed into arguments that the human gene pool should be improved by selective breeding policies, including incentives for reproduction for those of "good stock" and disincentives, such as compulsory sterilization, "euthanasia", and later, prenatal testing, birth control, and genetic engineering, for those of "bad stock". Another example of an extension of evolutionary theory that is now widely regarded as unwarranted is "Social Darwinism"; a term given to the 19th century Whig Malthusian theory developed by Herbert Spencer into ideas about "survival of the fittest" in commerce and human societies as a whole, and by others into claims that social inequality, racism, and imperialism were justified.[82]
See also
- For a more comprehensive list of topics, see Category:Evolution and Category:Evolutionary biology
|
|
|
Notes
- ↑ Desmond, Adrian; Moore, James (1991). Darwin. London: Michael Joseph, the Penguin Group. ISBN 0-7181-3430-3., [1] Darwin, C. R. ed. 1840. Fossil Mammalia Part 1 No. 4 of The zoology of the voyage of H.M.S. Beagle. By Richard Owen. P. 106]
- ↑ 2.0 2.1 Futuyma, Douglas J. (2005). Evolution. Sunderland, Massachusetts: Sinauer Associates, Inc. ISBN 0-87893-187-2.
- ↑ Gould, Stephen J. (2002). The Structure of Evolutionary Theory. Belknap Press. ISBN 0-674-00613-5.
- ↑ Lande, R.; Arnold, S.J. (1983). "The measurement of selection on correlated characters". Evolution 37: 1210–1226.
- ↑ Haldane, J.B.S. (1953). "The measurement of natural selection". Proceedings of the 9th International Congress of Genetics 1: 480–487.
- ↑ 6.0 6.1 Mechanisms: the processes of evolution. Understanding Evolution. University of California, Berkeley. Retrieved on 2006-07-14.
- ↑ Myers, PZ. "Ann Coulter: No evidence for evolution?", Pharyngula, scienceblogs.com, 2006-06-18. Retrieved on 2006-11-18.
- ↑ IAP Statement on the Teaching of Evolution Joint statement issued by the national science academies of 67 countries, including the United Kingdom's Royal Society (PDF file)
- ↑ From the American Association for the Advancement of Science, the world's largest general scientific society: 2006 Statement on the Teaching of Evolution (PDF file), AAAS Denounces Anti-Evolution Laws
- ↑ Stephen J. Gould (1997-06-12). Darwinian Fundamentalism. New York Review of Books. Retrieved on 2006-08-01.
- ↑ BEISSON, J. & SONNEBORN, T. M. (1965). Cytoplasmic inheritance of the organization of the cell cortex of Paramecium aurelia. Proc. natn. Acad Sci. U.S.A. 53, 275-282
- ↑ Snustad, P. and Simmons, A. 2000. Principles of Genetics, 2nd edition. John Wiley and Sons, Inc. New-York, p.20
- ↑ Carroll S.B,. Grenier J.K., Weatherbee S.D. (2005). From DNA to Diversity: Molecular Genetics and the Evolution of Animal Design. Second Edition. Oxford: Blackwell Publishing. ISBN 1-4051-1950-0.
- ↑ enmicro.pdf
- ↑ Pennisi_2003.pdf
- ↑ Dzidic S, Bedekovic V. (2003). "Horizontal gene transfer-emerging multidrug resistance in hospital bacteria". Acta pharmacologica Sinica 24 (6): 519-526.
- ↑ Andersson JO (2005). "Lateral gene transfer in eukaryotes". Cellular and molecular life sciences 62 (11): 1182-1197.
- ↑ Katz LA (2002). "Lateral gene transfers and the evolution of eukaryotes: theories and data". International journal of systematic and evolutionary microbiology 52 (5): 1893-1900.
- ↑ Evolutionary Theory by Peter Gogarten, Ph.D.
- ↑ Grant, P.R., and Grant, B.R. (1992). "Hybridization in bird species". Science 256 (4061): 193-7.
- ↑ Nowak et al. "Evolutionary dynamics of biological games"Science"'303'", 793-799 (2004), see also Nowak's book Evolutionary Dynamics
- ↑ see eg Axelrod & Hamilton, Science 211, 1390 (1981)
- ↑ There are now many papers on this, see eg Sachs Cooperation within and among species Journal of Evolutionary Biology 19, 1415 (2006)
- ↑ Nowak et al. Five Rules for the Evolution of Cooperation Science 314, 1560 (2006)
- ↑ Mayr, Ernst (1942). Systematics and the Origin of Species. NY: Columbia University Press.
- ↑ Hoskin et al (Oct 2005). "Reinforcement drives rapid allopatric speciation". Nature 437: 1353-1356.
- ↑ Savolainen et al (May 2006). "Sympatric speciation in palms on an oceanic island". Nature 441: 210-213.
- ↑ Barluenga et al (February 2006). "Sympatric speciation in Nicaraguan crater lake cichlid fish". Nature 439: 719-723.
- ↑ Feduccia, Alan (1996). The Origin and Evolution of Birds. New Haven: Yale University Press. ISBN 0-300-06460-8.
- ↑ Daeschler, Edward B., Shubin, Neil H., & Jenkins Jr, Farish A. (April 2006). "A Devonian tetrapod-like fish and the evolution of the tetrapod body plan". Nature 440: 757–763. DOI:10.1038/nature04639. Retrieved on 2006-07-14.
- ↑ Chimpanzee Sequencing and Analysis Consortium (2005). "Initial sequence of the chimpanzee genome and comparison with the human genome". Nature 437: 69–87.
- ↑ Britten, R.J. (2002). "Divergence between samples of chimpanzee and human DNA sequences is 5%, counting indels". Proc Natl Acad Sci U S A 99: 13633–13635.
- ↑ Schweitzer M.H. et al (2005). "Soft-tissue vessels and cellular preservation in Tyrannosaurus rex". Science 307 (5717): 1952-1955.
- ↑ Two sources: 'Genomic divergences between humans and other hominoids and the effective population size of the common ancestor of humans and chimpanzees'. and 'Quantitative Estimates of Sequence Divergence for Comparative Analysis of Mammalian Genomes' "[2] [3]"
- ↑ The picture labeled "Human Chromosome 2 and its analogs in the apes" in the article Comparison of the Human and Great Ape Chromosomes as Evidence for Common Ancestry shows how humans have a single chromosome which is two separate chromosomes in the nonhuman apes.
- ↑ The New York Times report Still Evolving, Human Genes Tell New Story, based on A Map of Recent Positive Selection in the Human Genome, states the International HapMap Project is "providing the strongest evidence yet that humans are still evolving" and details some of that evidence.
- ↑ Okamoto N, Inouye I. (2005). "A secondary symbiosis in progress". Science 310 (5746): 287.
- ↑ Okamoto N, Inouye I. (2006). "Hatena arenicola gen. et sp. nov., a Katablepharid Undergoing Probable Plastid Acquisition.". Protist Article in Print.
- ↑ Pseudogene evolution and natural selection for a compact genome. "[4]"
- ↑ Campbell & Reece (2002), Biology, Sixth Edition p. 446-447, ISBN 0-8053-6624-5
- ↑ Underhill et al, Nature Genetics 26, 358 - 361 (2000), "Y chromosome sequence variation and the history of human populations"
- ↑ Seielstad et al, Genome Research, Vol. 9, Issue 6, 558-567, June 1999, "A View of Modern Human Origins from Y Chromosome Microsatellite Variation"
- ↑ Evaluation of the Rate of Evolution in Natural Populations of Guppies (Poecilia reticulata) "[5]"
- ↑ Oklahoma State - Horizontal Gene Transfer: "Sequence comparisons suggest recent horizontal transfer of many genes among diverse species including across the boundaries of phylogenetic 'domains'. Thus determining the phylogenetic history of a species cannot be done conclusively by determining evolutionary trees for single genes."
- ↑ esalenctr.org
- ↑ In the years after Darwin's publication and fame, numerous "predecessors" to natural selection were discovered, such as William Charles Wells and Patrick Matthew, who had published unelaborated and undeveloped versions of similar theories earlier to little or no attention. Historians acknowledge that Darwin was the first to develop the theory rigorously and developed it independently. On Matthew, one historian of evolution has written that he "did suggest a basic idea of selection, but he did nothing to develop it; and he published it in the appendix to a book on the raising of trees for shipbuilding. No one took him seriously, and he played no role in the emergence of Darwinism. Simple priority is not enough to earn a thinker a place in the history of science: one has to develop the idea and convince others of its value to make a real contribution. Darwin's notebooks confirm that he drew no inspiration from Matthew or any of the other alleged precursors."
- ↑ Bowler, Peter J. (2003). Evolution: The History of an Idea. Berkeley: University of California Press, 158.
- ↑ http://www.life.umd.edu/emeritus/reveal/pbio/darwin/darwin02.html
- ↑ Darwin's proposal of a hereditary mechanism, pangenesis, lacked evidence and was ultimately rejected, being replaced by genetics.Darwin’s Theory of Pangenesis
- ↑ 50.0 50.1 Bowler, Peter J. (1989). The Mendelian Revolution: The Emergence of Hereditarian Concepts in Modern Science and Society. Baltimore: John Hopkins University Press.
- ↑ The convinced Mendelians, such as William Bateson and Charles Benedict Davenport, and biometricians, such as Walter Frank Raphael Weldon and Karl Pearson, became embroiled in a bitter debate, with Mendelians charging that the biometricians did not understand biology, and biometricians arguing that most biological traits exhibited continuous variation rather than the "jumps" expected by the early Mendelian theory. It is now recognized that the Mendelians were investigating Mendelian traits, those traits where existing variation is controlled by one gene and therefore is discrete, and the biometricians were investigating complex traits, where those traits were controlled by multiple genes, and the variation is therefore continuous.
- ↑ Resynthesizing evolutionary and developmental biology. Gilbert SF, Opitz JM, Raff RA. Developmental Biology 1996 Feb 1;173(2):357-72
- ↑ Rincon, Paul. "Evolution takes science honours", BBC News, 2005. Retrieved on 2006-07-16. According to the BBC, Colin Norman, news editor of Science, said "[S]cientists tend to take for granted that evolution underpins modern biology [...] Evolution is not just something that scientists study as an esoteric enterprise. It has very important implications for public health and for our understanding of who we are" and Dr. Mike Ritchie, of the school of biology at the University of St Andrews, UK said "The big recent development in evolutionary biology has obviously been the improved resolution in our understanding of genetics. Where people have found a gene they think is involved in speciation, I can now go and look how it has evolved in 12 different species of fly, because we've got the genomes of all these species available on the web."
- ↑ BBC Report on Biology Education in North America "In a study of 1,200 college freshmen, Professor Alters found that 45% of those who doubted the theory of evolution had specific misunderstandings about some of the science that has been used to support it."
- ↑ Constance Holden (1998). "SCIENCE EDUCATION: Academy Rallies Teachers on Evolution". Science 280 (5361): 194.
- ↑ Miller JD, Scott EC, Okamoto S. (2006). "Science communication. Public acceptance of evolution.". Science 313 (5788): 765-766.
- ↑ talkorigins Claim CB932: Evolution of degenerate forms
- ↑ M. Mitchell Waldrop (1992), Complexity, p. 296, Touchstone Book, ISBN 0-671-87234-6
- ↑ [6]
- ↑ talkorigins.org
- ↑ Richard Dawkins (1986), The Blind Watchmaker, p. 103, ISBN 0-393-31570-3
- ↑ Scientific American; Biology: Is the human race evolving or devolving?
- ↑ Anonymous. Claim CB901. The TalkOrigins Archive.
- ↑ Boxhorn, Joseph. Observed Instances of Speciation. Talk Origins Archive.
- ↑ Kramer EM, Donohue K. (2006). "Evolution. Traversing the adaptive landscape in snapdragons..". Science 313 (5789): 924-5.. PMID 16917048.
- ↑ Gibson G. (2005). "Evolution. The synthesis and evolution of a supermodel.journal = Science" 307 (5717): 1890-1.. PMID 15790836.
- ↑ Disorder — A Cracked Crutch For Supporting Entropy Discussions
- ↑ Does Life On Earth Violate the Second Law of Thermodynamics?
- ↑ Evolution and Information: The Nylon Bug. New Mexicans for Science and Reason.
- ↑ It wasn't a highly competent design because the bacteria weren't extracting a lot of energy from the process, just enough to get by. And it was based on a simply frame shift reading of a gene that had other uses. But with a simple frame shift of a gene that was already there, it could now "eat" nylon. Future mutations, perhaps point mutations inside that gene, could conceivably heighten the energy gain of the nylon decomp process, and allow the bacteria to truly feast and reproduce faster and more plentifully on just nylon, thus leading perhaps in time to an irreducibly complex arrangement between bacteria who live solely on nylon and a man-made fiber produced only by man. Darwinism or Directed Mutations?
- ↑ Prijambada I.D. et al (1995). "Emergence of nylon oligomer degradation enzymes in Pseudomonas aeruginosa PAO through experimental evolution". Applied and Environmental Microbiology 61 (5): 2020–2022.
- ↑ Negoro S et al (2005). "X-ray crystallographic analysis of 6-aminohexanoate-dimer hydrolase: molecular basis for the birth of a nylon oligomer-degrading enzyme". The Journal of Biological Chemistry 280 (47): 39644-39652.
- ↑ An overview of the philosophical, religious, and cosmological controversies by a philosopher who strongly supports evolution is: Daniel Dennett, Darwin's Dangerous Idea: Evolution and the Meanings of Life (New York: Simon & Schuster, 1995). On the scientific and social reception of evolution in the 19th and early 20th centuries, see: Peter J. Bowler, Evolution: The History of an Idea, 3rd. rev. edn. (Berkeley: University of California Press, 2003).
- ↑ 74.0 74.1 Burns, Edward M.; Philip Lee Ralph, Robert E. Lerner, Standish Meacham (1982). World Civilizations Their History and Their Culture. W.W. Norton & Company, 964-965. ISBN 0-393-95077-8.
- ↑ Huxley, Thomas H. (1902). Collected Essays, V. Kessinger Publishing. ISBN 978-1417973729.
- ↑ [7]
- ↑ The Rabbinical Council of America notes that significant Jewish authorities have maintained that evolutionary theory, properly understood, is not incompatible with belief in a Divine Creator, nor with the first 2 chapters of Genesis. [8]
- ↑ Aish HaTorah According to a possible reading of ancient commentators' description of God and nature, the world may be simultaneously young and old.
- ↑ Index to Creationist Claims, Claim CC150 edited by Mark Isaak. The TalkOrigins Archive, 2005
- ↑ Spergel, D. N.; et al. (2003). "First-Year Wilkinson Microwave Anisotropy Probe (WMAP) Observations: Determination of Cosmological Parameters". The Astrophysical Journal Supplement Series 148: 175—194. DOI:10.1086/377226.
- ↑ Wilde, S. A.; Valley J. W., Peck W. H. and Graham C. M. (2001). "Evidence from detrital zircons for the existence of continental crust and oceans on the Earth 4.4 Gyr ago". Nature 409: 175—178.
- ↑ On the history of eugenics and evolution, see Daniel Kevles, In the Name of Eugenics: Genetics and the Uses of Human Heredity (New York: Knopf, 1985).
General references
- Sean B. Carroll, 2005, Endless Forms Most Beautiful: The New Science of Evo Devo and the Making of the Animal Kingdom, W. W. Norton & Company. ISBN 0-393-06016-0
- Futuyma, D. (2005). Evolution. Sinauer Associates, Inc.. ISBN 0-87893-187-2.
- Natalia S. Gavrilova & Leonid A. Gavrilov, 2002,
- Gigerenzer, Gerd, et al., The empire of chance: how probability changed science and everyday life (New York: Cambridge University Press, 1989).
- Edward J. Larson, Evolution: The Remarkable History of a Scientific Theory (Modern Library Chronicles). Modern Library (May 4, 2004). ISBN 0-679-64288-9
- Garcia-Fernàndez, Jordi. Amphioxus: a peaceful anchovy fillet to illuminate Chordate Evolution. Int J Biol Sci (May, 2006).
- Mayr, Ernst. What Evolution Is. Basic Books (October, 2002). ISBN 0-465-04426-3
- Witzany, Guenther. Life:The Communicative Structure. Libri Books on Demand (October, 2000). ISBN 3-8311-0349-6
- Menand, Louis. 2001 The Metaphysical Club. New York: Farar, Straus and Giraux. ISBN 0-374-19963-9
- Williams, G.C. (1966). Adaptation and Natural Selection: A Critique of some Current Evolutionary Thought. Princeton, N.J.: Princeton University Press.
- Zimmer, Carl. Evolution: The Triumph of an Idea. Perennial (October 1, 2002). ISBN 0-06-095850-2
External links
- Understanding Evolution from University of California, Berkeley
- Evolution of Life Explained by Carl Sagan
- Natural Selection Explained by Carl Sagan
- National Academies Evolution Resources
- National Academy Press: Teaching About Evolution and the Nature of Science
- Evolution by Natural Selection, an introduction to the logic of evolution by natural selection
- Charles Darwin's writings
- A list of evidence for evolution compiled by PZ Myers.
- EvoWiki, a wiki whose goal is to promote general evolution education, and provide mainstream scientific responses to the arguments of antievolutionists
- Evolution (provided by PBS)
- Everything you wanted to know about evolution (provided by New Scientist)
- International Journal of Organic Evolution
- New England Complex Systems Institute
- Howstuffworks.com — How Evolution Works
- Synthetic Theory Of Evolution: An Introduction to Modern Evolutionary Concepts and Theories
- Evolution News from Genome News Network (GNN)
- Evolution for beginners
- RMCybernetics - AI Evolution can create emergent behavior in a computer program.
- NPR - Science Friday: links to museums, articles and books.
- "Evolution: Fact and Theory" by Richard E. Lenski
- Evolution by level Book reviews of books on evolution by knowledge level.
- Understanding Evolution: History, Theory, Evidence, and Implications Deals heavily with the history of evolutionary thought
- Becoming Human - Journey through the story of human evolution
- Evolution & ID Timeline Focuses on major historical and recent events in the scientific and political debate
- Timeline of Evolutionary Thought The life and work of notable people who have contributed to evolutionary thought
- Talk.Origins Archive — see also talk.origins
- Not Just a Theory Discredits the assertion that evolution is "just a theory", with an explanation of the meaning of the word 'theory' in a scientific context.
- Dispelling the Top Ten Myths About Evolution Endorsed by Ann Druyan and the American Association for the Advancement of Science: introduces the top ten common myths and misunderstandings about evolution and includes a sample chapter, "Survival of the Fittest".
Evolution simulators
- Isolated species evolves to interact more efficiently with its environment (java applet)
- Evolution in a predator-prey relationship (java applet)
- Watch small creatures evolve into more efficient swimmers
- Blind Watchmaker Applet (java)
Basic topics in evolutionary biology
|
|---|
| Evidence of evolution |
| Processes of evolution: adaptation - macroevolution - microevolution - speciation |
| Population genetic mechanisms: selection - genetic drift - gene flow - mutation |
| Evo-devo concepts: phenotypic plasticity - canalisation - modularity |
| Modes of evolution: anagenesis - catagenesis - cladogenesis |
| History: History of evolutionary thought - Charles Darwin - The Origin of Species - modern evolutionary synthesis |
| Other subfields: ecological genetics - human evolution - molecular evolution - phylogenetics - systematics |
| List of evolutionary biology topics | Timeline of evolution |
Topics in population genetics
|
|---|
| Key concepts: Hardy-Weinberg law | genetic linkage | linkage disequilibrium | Fisher's fundamental theorem | neutral theory |
| Selection: natural | sexual | artificial | ecological |
| Effects of selection on genomic variation: genetic hitchhiking | background selection |
| Genetic drift: small population size | population bottleneck | founder effect | coalescence |
| Founders: R.A. Fisher | J.B.S. Haldane | Sewall Wright |
| Related topics: evolution | microevolution | evolutionary game theory | fitness landscape | genetic genealogy |
| List of evolutionary biology topics |